Introduction
The eyes are complex sense organs that have evolved from primitive light-sensitive spots on the surface of invertebrates. Within its protective casing, each eye has a layer of receptors, a lens system that focuses light on these receptors, and a system of nerves that conducts impulses from the receptors to the brain.
Anatomic considerations
The outer protective layer of the eyeball, the sclera, is modified anteriorly to form the transparent cornea, through which light rays enter the eye. Inside the sclera is the choroid, a layer that contains many of the blood vessels that nourish the structures in the eyeball. Lining the posterior two-thirds of the choroid is the retina, the neural tissue containing the receptor cells.
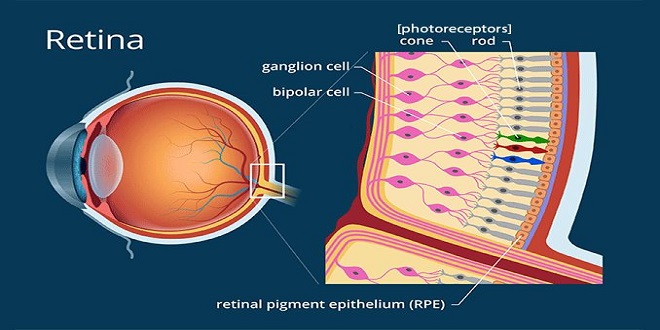
Retina
The retina extends anteriorly almost to the culinary body. It is organized in 10 layers and contains the rods and cones, which are the visual receptors, plus four types of neurons: bipolar cells, ganglion cells, horizontal cells, and murine cells. There are many different synaptic transmitters. The rods and cones, which are next to the choroid, synapse with bipolar cells, and the bipolar cell’s synapse with ganglion cells. About 12 different types of bipolar cells occur, based on morphology and function. The axons of the ganglion cells converge and leave the eye as the optic nerve. Horizontal cells connect receptor cells to the other receptor cells in the outer plexiform layer. Marine cells connect ganglion cells to one another in the inner plexiform layer via processes of varying length and patterns. At least 29 types of marine cells have been described on the basis of their connections. Gap junctions also connect retinal neurons to one another, and the permeability of these gap junctions is regulated
Receptors
Each rod and cone is divided into an outer segment, an inner segment that includes a nuclear region, and a synaptic zone. The outer segments are modified cilia and are made up of regular stacks of flattened scales or disks composed of the membrane. These scales and disks contain the photosensitive compounds that react to light, initiating action potentials in the visual pathways. The inner segments are rich in mitochondria. The rods are named for the thin, rod-like appearance of their outer segments. Cones generally have thick inner segments and conical outer segments, although their morphology varies from place to place in the retina. In cones, the scales are formed in the outer segments by inholdings of the cell membrane, but in rods, the disks are separated from the cell membrane.
The image-forming mechanism
The eyes convert energy in the visible spectrum into action potentials in the optic nerve. The wavelengths of visible light range from approximately 397–723 nm. The images of objects in the environment are focused on the retina. The light rays striking the retina generate potentials in the rods and cones. Impulses initiated in the retina are conducted to the cerebral cortex, where they produce the sensation of vision
Common defects of the image-forming mechanism
In some individuals, the eyeball is shorter than normal and the parallel rays of light are brought to a focus behind the retina. This abnormality is called hyperopia or farsightedness (Figure 12–9). Sustained accommodation, even when viewing distant objects, can partially compensate for the defect, but the prolonged muscular effort is tiring and may cause headaches and blurring of vision.
The photoreceptor mechanism
The potential changes that initiate action potentials in the retina are generated by the action of light on photosensitive compounds in the rods and cones. When light is absorbed by these substances, their structure changes and this triggers a sequence of events that initiates neural activity
The eye is unique in that the receptor potentials of the photoreceptors and the electrical responses of most of the other neural elements in the retina are local, graded potentials, and it is only in the ganglion cells that all-or-none action potentials transmitted over appreciable distances are generated.
Ionic basis of photoreceptor potentials
Na+ channels in the outer segments of the rods and cones are open in the dark, so current flows from the inner to the outer segment. Current also flows to the synaptic ending of the photoreceptor. The Na+–K+ pump in the inner segment maintains ionic equilibrium. The release of the synaptic transmitter is steady in the dark. When light strikes the outer segment, the reactions that are initiated close some of the Na+ channels, and the result is a hyperpolarizing receptor potential. The hyperpolarization reduces the release of synaptic transmitters, and this generates a signal in the bipolar cells that ultimately leads to action potentials in ganglion cells. The action potentials are transmitted to the brain.
Cone pigments
Primates have three different kinds of cones. These receptors subserve color vision and respond maximally to light at wavelengths of 440, 535, and 565 nm. Each contains retinene1 and a posing. The posing resembles rhodopsin and spans the cone membrane seven times but has a characteristic structure in each type of cone. The cell membrane of cones is invigilated to form the scales, but the cones have no separate intracellular disks like those in rods. The details of the responses of cones to light are probably similar to those in rods.
Resynthesis of cyclic GMP
Light reduces the concentration of Ca2+ as well as that of Na+ in photoreceptors. The resulting decrease in Ca2+ concentration activates gaunt cycles, which generates more cGMP. It also inhibits the light-activated phosphodiesterase. Both actions speed recovery, restoring the Na+ channels to their open position.
Processing of visual information in the retina
In a sense, the processing of visual information in the retina involves the formation of three images. The first image, formed by the action of light on the photoreceptors, is changed to a second image in the bipolar cells, and this in turn is converted to a third image in the ganglion cells. In the formation of the second image, the signal is altered by the horizontal cells, and in the formation of the third, it is altered by the murine cells. There is little change in the impulse pattern in the lateral geniculate bodies, so the third image reaches the occipital cortex.
Responses in the visual pathways & cortex
The axons of retinal ganglion cells project a detailed spatial representation of the retina on the lateral geniculate body. Each geniculate body contains six well-defined layers. Layers 3–6 have small cells and are called parvocellular, whereas layers 1 and 2 have large cells and are called magnocellular. On each side, layers 1, 4, and 6 receive input from the contralateral eye, whereas layers 2, 3, and 5 receive input from the ipsilateral eye. In each layer, there is a precise point-for-point representation of the retina, and all six layers are in the register so that along a line perpendicular to the layers, the receptive fields of the cells in each layer are almost identical. It is worth noting that only 10–20% of the input to the lateral geniculate nucleus comes from the retina. Major inputs also occur from the visual cortex and other brain regions. The feedback pathway from the visual cortex has been shown to be involved in visual processing related to the perception of orientation and motion.
Summary
The major parts of the eye are the sclera (protective covering), the cornea (transfer light rays), choroids (nourishment), retina (receptor cells), lens, and iris. The visual pathway is from the rods and cones to bipolar cells to ganglion cells then via the optic tract to the thalamic lateral geniculate body to the occipital lobe of the cerebral cortex. The fibers from each nasal hemiretina decussate in the optic chiasm; the fibers from the nasal half of one retina and the temporal half of the other synapse on the cells whose axons form the geniculo calcarine tract.